Von Dr. Andreas Müller
Warmblüter und Kaltblüter
Die Mehrzahl aller Tierarten auf der Erde kann die eigene Körpertemperatur nicht wesentlich über die Umgebungstemperatur steigern. Das heißt, wenn es kalt wird, sind diese Tiere mehr oder weniger zur Regungslosigkeit verurteilt. Diese Arten nennt man umgangssprachlich „Kaltblüter“ oder „Wechselwarme“ (wissenschaftlich: Poikilotherme) und zu ihnen gehören alle Wirbellosen, Fische, Amphibien und Reptilien. Zu den „Warmblütern“ oder Gleichwarmen“ (wissenschaftlich: Homoiotherme) gehören Vögel und Säugetiere, da sie ihre Körpertemperatur auf einem etwa gleichen Niveau regulieren können. Allerdings kostet diese Wärmeregulation viel Energie, die durch Nahrungszufuhr gewonnen werden muss. Deshalb benötigen Säugetiere, zu denen auch die Fledermäuse gehören, deutlich mehr Nahrung für ihren Stoffwechsel als beispielsweise Eidechsen. Durch einen Gefieder- oder Fellwechsel oder durch eine verringerte Durchblutung der Haut können Warmblüter ihre Wärmeisolation verbessern und dadurch auch ihre Energiebilanz.
Infolge ihres Körperbaus und der Tatsache, dass sie als einzige Säugetiere fliegen können, sind Fledermäuse besonders zum Energiesparen verpflichtet und haben von allen Säugetieren die größte Anzahl an Temperaturregulationsmechanismen. Aufgrund ihrer stark durchbluteten Flughäute ist ihre Körperoberfläche im Verhältnis zur Körpermasse viel größer als bei beispielsweise gleichschweren Mäusen oder Spitzmäusen. Aufgrund ihrer Größe können Flughunde oder auch größere Fledermäuse in den Tropen ihre Körpertemperatur Tag und Nacht und auch während des Fluges oder des Ruhens auf gleichem Niveau halten. Unsere heimischen, deutlich kleineren Fledermäuse können dies nicht so einfach.
Tagesschlaflethargie oder Torpor
In unseren gemäßigten Klimazonen zeigen Fledermäuse deshalb auch einen Unterschied in der Wärmeregulation während der Wach- und Ruhephasen. Wie der Fledermausforscher Martin Eisentraut feststellte, zeigten Fledermäuse, die in ihren Sommerquartieren am Tage untersucht wurden, zu niedrige Körpertemperaturen (etwa 16 bis 18 °C), um sich noch groß bewegen zu können oder spontan aufzufliegen, was Eisentraut als „Tagesschlaflethargie“ bezeichnete. Heute wird dieser Zustand als Torpor bezeichnet. In der Nacht stieg dann die Körpertemperatur wieder auf 35 bis 42°C an, also auf eine gute Betriebstemperatur zum Fliegen.
Dabei hängt die Tagesschlaftemperatur der Fledermaus von der Umgebungstemperatur ab und je tiefer die Körpertemperatur ist, desto mehr ist die Bewegungsfähigkeit der Fledermaus eingeschränkt. Dieser Torpor kann recht schnell unterbrochen werden, da sich die Fledermäuse in etwa 30 Minuten wieder auf Betriebstemperatur aufwärmen können. In Kolonien von Fledermäusen wurde beobachtet, dass es an kühlen Tagen (um die 11°C) aktive und lethargische Tiere gab, alle Männchen waren in Torpor, während die Weibchen aktiv ihre Jungen säugten. Als die Temperatur weiter sank, gingen auch die Weibchen und Jungtiere in Torpor. Ein Topor ist verbunden mit der Reduktion der Körpertemperatur, dem Abfall der Sauerstoffaufnahme (reduzierte Atmung), der Verringerung der Herzschlagfrequenz und einem reduzierten Blutfluss in die Extremitäten, deren Blutgefäße sich zusammenziehen.
Winterschlaf
Der Winterschlaf ist ist ein besonderer Zustand eines Torpors, der tage-, wochen-, oder monatelang (bis zu 7 Monate) andauern kann. Er bricht nicht spontan über die Fledermaus ein, sondern ist ein kontrollierter und jahresrhythmischer Prozess. In unseren gemäßigten Breitengraden halten Fledermäuse Winterschlaf, da im Winter das Nahrungsangebot an Insekten und anderen Gliedertieren sehr gering ist. Deshalb haben sie nur zwei Alternativen: Abwandern in wärmere Gegenden mit größeren Nahrungsangebot oder in den Winterschlaf gehen und die Fettreserven aus dem Sommer/Herbst aufbrauchen. Viele Fledermausarten überstehen den Winter in einer Dauerlethargie, wobei ihre Körpertemperatur noch unter die erreichten Werte bei der Tagesschlaflethargie fällt, manche Fledermäuse können sogar bis zum Gefrierpunkt (0°C) abkühlen.
Dann sind die Fledermäuse fast völlig starr und nur zu Reflexbewegungen fähig. Nur wenn genügend Energievorräte vorhanden sind, behält eine Fledermaus auch die Kontrolle über ihre Körpertemperaturregelung, sollten diese aufgebraucht sein, unterkühlt sie und stirbt. Damit so wenig wie möglich Energie verbraucht wird, wird die Atmung heruntergeregelt. Einzelne Atemzüge können durch minutenlange Atempausen unterbrochen sein, da der Sauerstoffverbrauch im tiefen Winterschlaf extrem niedrig ist – zwischen 0,02 – 0,07 Milliliter je Gramm und Stunde, verglichen mit 3 Milliliter je Gramm und Stunde im aktiven Zustand. Außerdem sinkt die Herzfrequenz von 400 Schläge pro Minute im aktiven Tier auf 15 – 20 Schläge pro Minute ab, was etwa einem Vierzigstel des höchsten Wertes im Wachzustand entspricht. So kommt eine ruhende Fledermaus mit ihren Reserven etwa 100 Mal länger aus als ein aktives Tier.
Als Energiereserve wird dann in erster Linie Fett verbrannt, hauptsächlich das sogenannte „braune Fettgewebe“, das wie ein Polsterkissen zwischen den Schulterblättern und an den Flanken der Fledermaus liegt. Bei guter Witterung fressen sich Fledermäuse bis in den November ihre Fettreserven an und durch eine häufigere tiefe Tageslethargie erhalten sie diese Reserven auch, da ihr Energieverbrauch am Tage niedrig ist. Vor der Winterruhe wiegen die Fledermäuse aufgrund dieses Fettpolsters 20-30% mehr, als in den Sommermonaten und mit diesem Polster müssen die Tiere dann bis zu sieben Monate auskommen. Während des Winterschlafes müssen die Tiere weder Harn noch Kot lassen.
Aufwachphase
Der Aufwachvorgang hängt wesentlich von der Umgebungstemperatur ab. Er beginnt mit einer erhöhten Herzschlagfrequenz, dann folgt die Steigerung der Atemfrequenz und dem gesteigerten Blutstrom in den vorderen Körper. In der ersten Phase des Aufwachens, der Aufheizphase, stammen 80% der Energie aus dem braunen Fettgewebe und dies wird verbrannt, bis der Körper auf 15 °C aufgewärmt ist.
Zuerst werden Herz, Leber und Kopf erwärmt, später der restliche Körper. In der zweiten Phase wird das Muskelzittern als zusätzliche Wärmequelle eingesetzt und der Körper heizt sich weiter auf. Die Aufwachdauer hängt von der Temperaturspanne ab, die durchlaufen werden muss, um den Bereich der normalen Körpertemperatur zu erreichen. Die Aufwachphase ist abgeschlossen, wenn das Tier flugfähig ist, was zwischen 30 – 50 min dauern kann, je nach Umgebungstemperatur im Winterquartier.
Winterquartiere
Im November suchen die Fledermäuse, abhängig von der Umgebungstemperatur und dem Nahrungsangebot, ihre Winterschlafplätze auf. Haben sie diese gefunden, senken sie langsam ihre Atem- und Herzfrequenz herab und gehen in Lethargie. Der Körper kühlt nach und nach auf eine Temperatur ab, die etwa der Umgebungstemperatur entspricht. Dabei werden gezielt Quartiere gesucht, die den geringen Energieverbrauch unterstützen, also relativ konstante Temperaturen haben. Eine Art Kältewarnung schützt vor Fledermäuse vor der zu großen Auskühlung, wenn etwa größere Kälte in das Winterquartier einbricht.
Als Folge produziert die Fledermaus dann vermehrt Wärme – Herzfrequenz und Atemfrequenz steigen leicht an, damit das Tier nicht erfriert. Diese Kältewarnung reagiert bis zum Erwachen im Frühjahr. Gehen die Fettreserven zur Neige, können Fledermäuse auch aufwachen, um im Winter auf Nahrungssuche zu fliegen – wenige Insekten, wie etwa der Frostspanner, ein Schmetterling, fliegen auch in Winternächten.
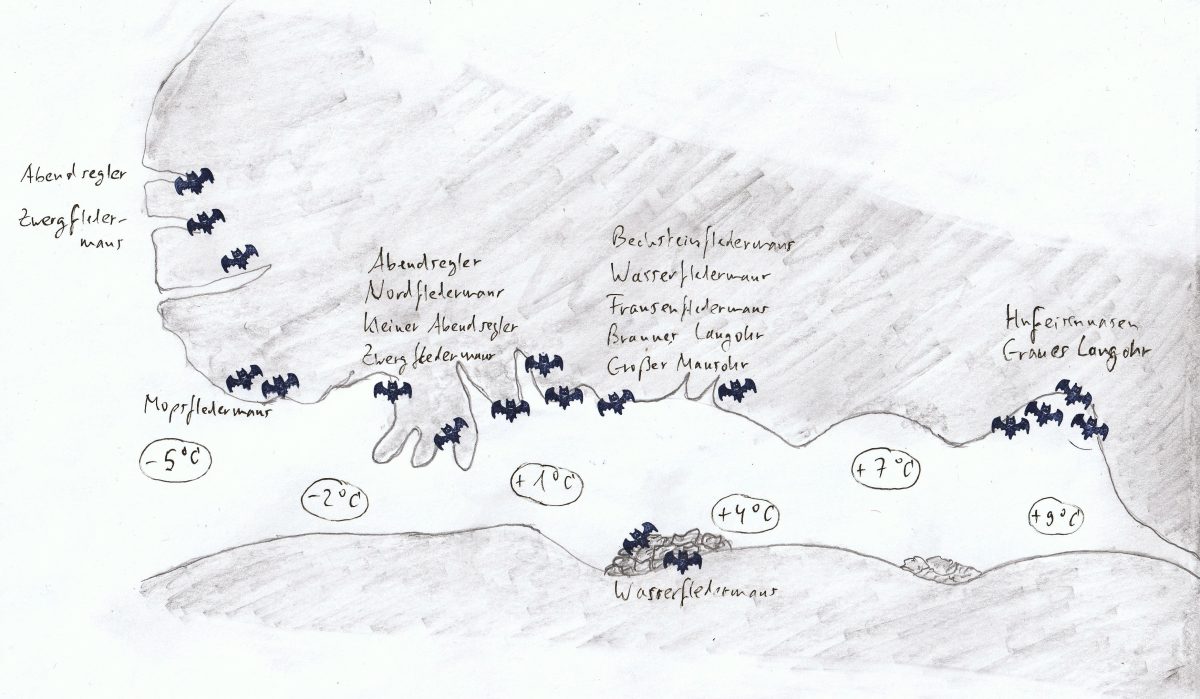
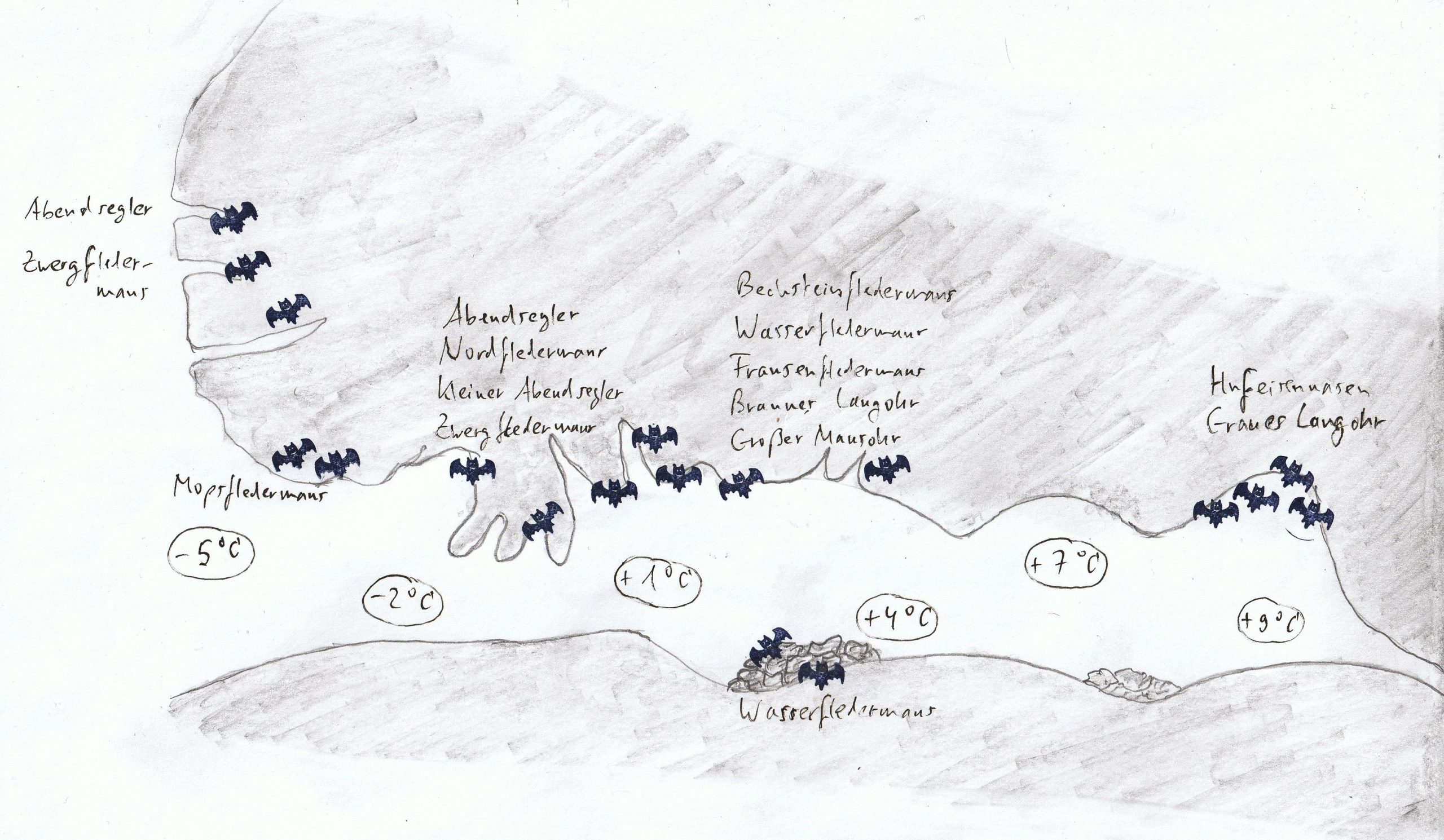
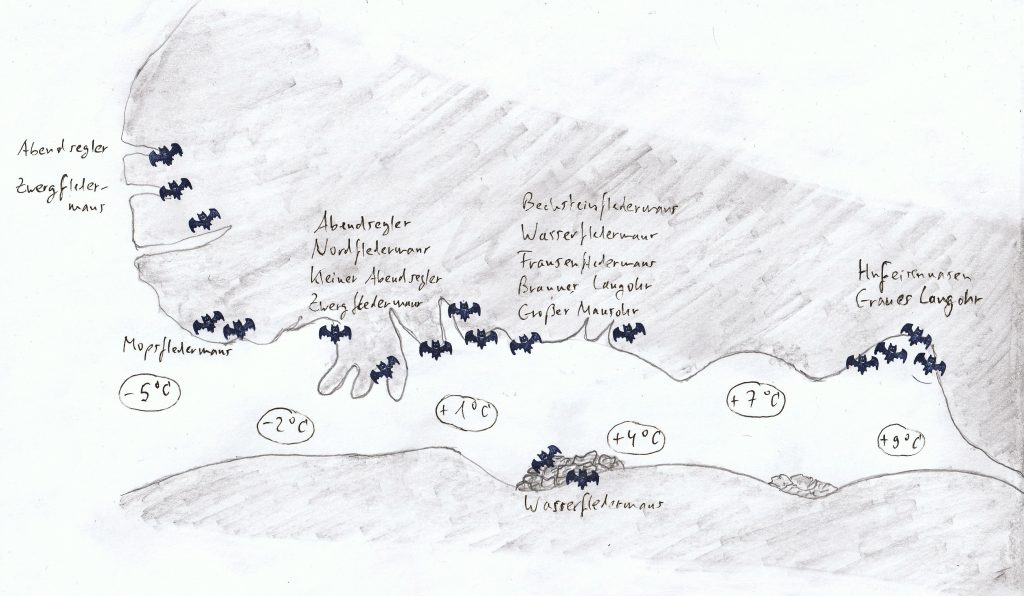
Je nach Winterquartier unterscheidet man zwischen Baum- und Felsfledermäusen. Erstere bevorzugen Baumhöhlen und Rindenspalten, letztere Höhlen, Gebäude, Keller, Bunker und Dachstühle. Da Höhlen und Gebäude konstantere Temperaturen und Bedingungen, verglichen mit Baumhöhlen bieten, sind Baumfledermäuse robuster in Bezug auf Kälte und Luftfeuchtigkeit. Typische Felsfledermäuse sind Mausohren und ihre Verwandten (Gattung Myotis), Langohren (Gattung Plecotus), die Mopsfledermaus (Barbastella barbastellus), sowie die Hufeisennasen (GattungRhinolophus).
Die einzelnen Arten bevorzugen bestimmte Plätze in einer Höhle: Zwergfledermäuse (Pipistrellus pipistrellus) und Abendsegler (Gattung Nyctalus) überwintern an den kühlsten Stellen in der Nähe des Höhleneingangs, Langohren und Mausohren haben die breiteste Toleranz und überwintern an Stellen in der Mitte der Höhle, Bartfledermäuse und Wasserfledermäuse in der Mitte einer Höhle, während Hufeisennasen die geringste Temperaturtoleranz haben und deshalb ganz weit im Inneren einer Höhle hängen.
Dabei suchen manche Arten, wie Zwergfledermäuse, eher Spalten und Ritzen in Felsen, die am Eingang einer Höhle liegen und nachts extrem kalt sind, sich aber tagsüber durch die Sonne etwas erwärmen. Andere Arten, wie die Langohren haben ein ganz anderes Problem, da ihre abstehenden großen Ohren im Winter einfrieren würden. Deshalb klappen sie diese nach hinten und verstecken sie unter den Flügeln. Nur der Tragus (der Ohrdeckel) ist dann noch zu sehen. Wasserfledermäuse überwintern gerne in Spalten innerhalb der Höhle oder unter Geröll am Boden, während Hufeisennasen freischwebend, in ihre Flügel eingewickelt, an der Decke hängen und somit Temperatur- und Feuchtigkeitsveränderungen direkt ausgesetzt sind. Wenn sich die Temperatur oder Luftfeuchtigkeit im Winter ändert, wandern Fledermäuse entweder weiter ins Innere der Höhle oder näher zum Eingang.
Typische Baumfledermäuse sind die Abendsegler (Gattung Nyctalus), die gerne als Winterquartier, Baumhöhlen besetzen, wo sie Cluster (Gemeinschaften aus mehreren Tieren) bilden, die sich gegenseitig wärmen. Solche Cluster können auch aus mehreren Fledermausarten bestehen. Im Frühjahr oder oft auch im Winter werden solche Baumhöhlen schnell erwärmt, wodurch die Fledermäuse früher aktiv werden und man an warmen Winterabenden Abendsegler fliegen sehen kann. Bei Baumpflegemaßnahmen im Winter in Parks oder Wäldern empfiehlt es sich, zu fällende Bäume auf mögliche Höhlen, in denen Fledermäuse überwintern könnten, zu untersuchen.
Quartiere von Fledermausarten in Höhlen (verändert nach Nagel in Siemers & Nill 2000)
Störungen der Winterquartiere durch den Menschen können sich als besonders gefährlich auswirken, weil bereits das Anleuchten mit der Taschenlampe, das Blitzlicht eines Fotoapparates oder auch der längere Aufenthalt in der Nähe der Fledermäuse als Weckreiz für ein Tier wirken kann. Die Fledermäuse erwachen und verbrauchen dabei wertvolle Energiereserven, die ihnen dann im Frühjahr für den Aufwachprozess fehlen. Deshalb sollten Fledermäuse so wenig wie möglich (oder nötig) in ihren Winterquartieren gestört werden, egal ob dies Höhlen, Dachstühle, Bunker, Keller oder Baumstämme sind.
Quellen
Altringham 2011: Bats. From Evolution to Conservation – 2011. Oxford University Press, Oxford
Eisentraut 1957: Aus dem Leben der Fledermäuse und Flughunde. Gustav Fischer Verlag, Jena
Gebhard 1997: Fledermäuse. Birkhäuser Verlag, Basel
Neuweiler 1993: Biologie der Fledermäuse. Georg Thieme Verlag, Stuttgart
Richarz & Limbrunner 2003: Fledermäuse. Fliegende Kobolde der Nacht. Franckh-Kosmos Verlags-GmbH & Co., Stuttgart
Siemers & Nill 2000: Fledermäuse. Das Praxisbuch. BLV-Verlag, München
© Text und Zeichnung Dr. Andreas Müller, Düsseldorf