

Dr. Andreas Müller, Düsseldorf
Anders als etwa bei Dinosauriern, existieren von Fledertieren (Flughunden und Fledermäusen) nur wenige gut erhaltene fossilisierte Skelette – hier ist vor allem die Grube Messel bei Darmstadt als Fundort zu nennen. Da Fledertiere in der Erdgeschichte schon immer klein waren (mit Ausnahme einiger Flughunde) und aufgrund ihrer Fähigkeit zum Flug dünne, leichte und röhrenartige Knochen haben, sind oft nur fossilisierte Zähne vom Skelett übriggeblieben, nach denen Paläontologen eine neue Art beschreiben können. So sind die meisten fossilen Fledertiere nur durch ihre Zähne belegt und beschrieben. Im Weiteren sollen deshalb nur die durch Skelette oder Teilskelette dokumentierten Fledertiere betrachtet werden.
Ursprung der Fledertiere
Den Ursprung der Fledertiere vermuten Forscher in kleinen Spitzmaus-ähnlichen, baumbewohnenden Säugetieren, die von Ast zu Ast sprangen und auf der Jagd nach Gliedertieren (Insekten und Spinnen) oder kleinen Wirbeltieren waren. Durch das Springen und Gleiten im Geäst entwickelte sich bei diesen Vorfahren nach und nach eine Flughaut, ähnlich wie bei heutigen Flughörnchen. Aus den Vordergliedmaßen entwickelte sich dann ein Flügel, während sich die Hintergliedmaßen verkürzten und für eine Bewegung am Boden immer ungeeigneter wurden. So bewegen sich die meisten heutigen Fledermäuse auch eher unbeholfen am Boden, während ihr Flug ganz anders aussieht, der zu den am besten entwickelten Formen in der Tierwelt gehört. Leider ist bis heute keiner der Vorfahren fossil überliefert, die ältesten Fledermausfossilien zeigen schon Flügel für einen aktiven Flug (nicht nur für einen Gleitflug).
Fossile Fledermausfunde aus Wyoming, USA
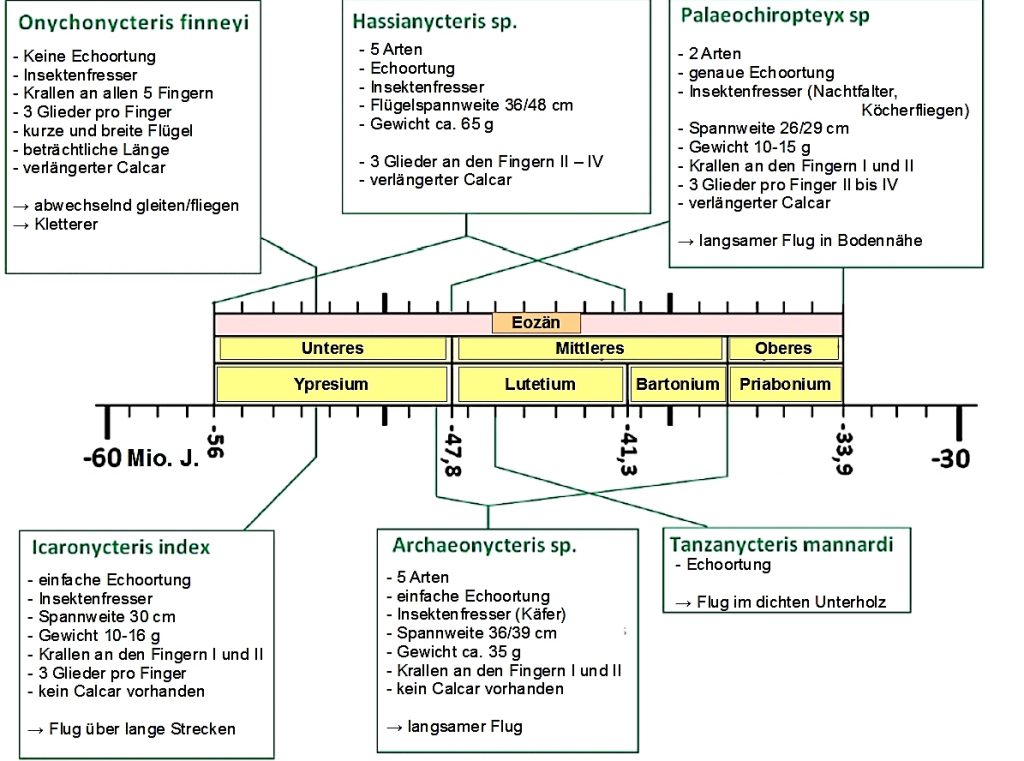
Die älteste, fossil überlieferte Fledermaus stammt aus dem unteren Eozän (vor etwa 52 Millionen Jahren), gefunden in der Green River Formation in Wyoming, USA und wurde Onychonycteris finneyi genannt. Onychonycteris besitzt einen Flügel mit sehr kurzen Fingergliedern, die Hinterbeinproportionen sind etwas primitiver und länger, als bei heute lebenden Fledermäusen und ein langer Calcar ist vorhanden, was auf eine breitere Schwanzmembran hindeutet. Der Calcar ist ein knorpeliger Sporn bei Fledermäusen, der zur Stützung der Flughaut dient. Er ist an den Hinterbeinen ausgebildet und spannt die Schwanzflughaut zwischen Hinterbein und Schwanz der Tiere. Alle diese Strukturen deuten darauf hin, dass Onychonycteris mit ihren breiten Flügeln zu einem wellenförmigen, gleitend-flatternden Flug fähig war, der jedoch primitiver war als derjenige der heutigen Fledermäuse. Die fossilen Knochen des Ohrs deuten zudem darauf hin, dass Onychonycteris keine Echoortung, wie alle heutigen Fledermäuse besessen hat. Die Proportionen der Gliedmaßen und das Vorhandensein von Krallen an allen Finger deuten darauf hin, dass Onychonycteris ein wendiger Kletterer gewesen sein könnte, der sich quadruped (vierfüßig) fortbewegte und an Ästen hängend lebte.
Außerdem wurde in der Green River Formation eine weitere, gut erhaltene Fledermausart aus dem frühen Eozän (50 Mio. J.) entdeckt, die den Namen Icaronycteryx index bekam. Icaronycteryx war schon weiterentwickelt als Onychonycteris und verwandt mit den, in der Grube Messel gefundenen Archaeonycteris–Arten (siehe unten), zeigte aber noch eine zusätzliche Kralle am zweiten Finger, die bei heutigen Fledermäusen fehlt. Außerdem hatte Icaronycteryx, wie die Fledermäuse der Grube Messel schon eine Echoortung entwickelt.
Fossile Fledermausfunde aus der Grube Messel, Deutschland
Die weltweit älteste durch vollständige Skelette bekannte Fledermausgesellschaft stammt aus der Grube Messel bei Darmstadt. Sie wird auf ein Alter von 47 Millionen Jahren (frühes, mittleres Eozän) datiert. Die sieben bekannten Fledermausarten besaßen schon so unterschiedliche Flügelformen, dass man sie den Flug- und Jagdbiotopen heute existierender Fledermausgesellschaften zuordnen kann. Auch die Variationsbreite der Flügelformen und die Flächenbelastung deckte bereits quantitativ die Variationsbreite heutiger Fledermäuse ab. Diese Gesellschaft bestand aus Arten der Gattungen Archaeonycteris, Palaeochiropteryx, Hassianycteris und Tachypteron.
Die Gattung Archaeonycteris ist eng mit der in Wyoming ausgegrabenen Fledermausart Icaronycteryx index verwandt und hatte eine Flügelform mit mittlerer Flächenbelastung, was auf einen unspezialisierten Flug in mittlerer Flughöhe hindeutet. Die Flügelspannweite betrug bei Archaeonycteris trigodon 36,9 cm und bei Archaeonycteris pollex 39 cm. Wie Icaronycteryx zeigte auchArchaeonycteris neben der Daumenkralle eine weitere Kralle am Zeigefinger.
Die Arten der Gattung Palaeochiropteryxwaren breitflügelige Fledermäuse mit zierlichem Körperbau, weshalb die Flügel nur mit einem geringen Gewicht belastet waren. Sie hatten einen langsamen, wendigen Flug dicht über dem Boden oder nahe am Blattwerk der Bäume und Sträucher. Palaeochiropteryx tupaidon hatte eine Flügelspannweite von 26 cm undPalaeochiropteryx spiegeli von 29 cm.
Palaeochiropteryx tupaidon – Grube Messel
Zwei in ihren Zahnmerkmalen fortschrittliche Fledermausarten waren Hassianycteris messelenis und Hassianycteris magna. Sie besaßen eine schmale Flügelform und waren schnelle und hochfliegende Fledermäuse, die um die Baumwipfel flogen und eine, mit 39 cm bzw. 48 cm große Flügelspannweite besaßen.
Alle oben genannten Fledermausgattungen sind bereits ausgestorben, während die vierte in der Grube Messel gefundene Art Tachypteron franzeni der heute noch existierenden Famile Emballonuridae (Glattnasen-Freischwänze oder Sackflügelfledermäuse) angehört. Tachypteron hatte eine schmale Flügelform und bewegte sich schnell und hochfliegend im freien Luftraum. Ihre Flügelspannweite betrug 34,9 cm. Alle diese Fledermausarten aus dem Eozän besetzten unterschiedliche ökologische Nischen, wie das in heute zusammenlebenden Fledermausgesellschaften auch beobachtet werden kann (siehe Abbildung unten).
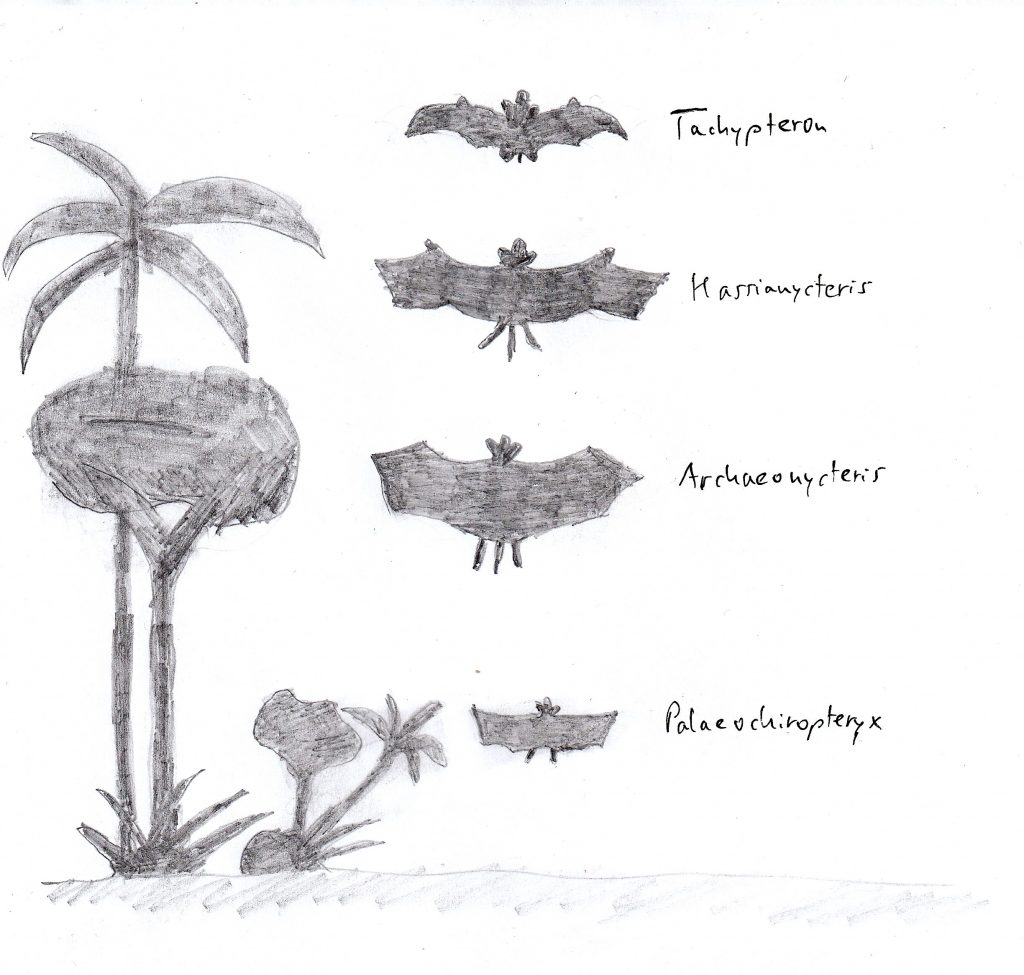
Fledermausgesellschaft in der Grube Messel vor 47 Millionen Jahren (verändert nach Habersetzer et al. 2008)
Fossile Fledermausfunde aus Tansania, Afrika
Die Fledermaus Tanzanycteris mannardi wurde 2003 aus Tansania in Afrika beschrieben und ist durch ein teilweise erhaltenes Skelett bekannt. Es besteht aus der vorderen Hälfte des Skeletts einschließlich Schädel, Unterkiefer, Wirbelsäule, Wirbelsäule, beiden Schultergürteln und der linken Speiche. Die Handgelenke, Flügelelemente, der größte Teil des Beckens und die hinteren Gliedmaßen fehlen. Tanzanycteris lebte vor 46 Mio. J. und besaß schon Echoortung. Am ehesten verwandt ist sie mit Hassianycteris aus der Grube Messel.

Übersicht fossile Fledermäuse
Molekulare Phylogenie der Fledertiere
Emma Teeling und Kollegen entwickelten 2005 eine Phylogenie (Stammesgeschichtliche Entwicklung) der heute existierenden Fledertiere basierend auf molekularbiologischen Methoden, die im unteren Eozän vor 52 – 50 Mio. Jahren begann zu einer Zeit, wo die Temperatur angestiegen ist, die Anzahl der Pflanzenarten und auch die der Insekten deutlich zunahm, alles günstige Bedingungen für die Entwicklung der Fledermäuse.
Die Forscher vermuten den Ursprung für drei Hauptlinien der Fledermäuse in Laurasia vor etwa 65 Mio. Jahren, möglicherweise in Nordamerika, während die vierte Hauptlinie ihren Ursprung in Gondwana, wahrscheinlich Südamerika hat. Kurz ein wenig Erdgeschichte: Im Perm (vor 298 – 252 Mio. J.) bestand die Erde aus einem Superkontinent, der Pangäa genannt wurde. Dieser zerfiel in der Trias (vor 252 – 201 Mio. J.) langsam in zwei Kontinente, Laurasia und Gondwana. Laurasia bestand aus den heutigen Kontinenten Asien und Nordamerika – Gondwana aus Südamerika, Afrika, Europa und Australien.
Aufgrund der phylogenetischen Untersuchungen von Teeling und Kollegen zeigte sich, dass die bisherige Unterscheidung in Mikrochiroptera (Fledermäuse) und Makrochiroptera (Flughunde) nicht korrekt war, da die Hufeisennnasen (Familie Rhinolophidae) und Großblattnasen (Fam. Megadermatidae) näher mit den Flughunden (Fam. Pteropodidae) verwandt ist als mit allen anderen Fledermäusen. Die Fledertiere wurden deshalb in die Yinpterochiroptera (Flughunde und Hufeisennasenartige) und in die Yangochiroptera (Fledermäuse) eingeteilt. Die ausgestorbenen Gattungen und Familien der Fledermäuse wurden in diesen Stammbaum eingegliedert.
Hypothesen zur Evolution der Fledertiere
Da ein Vorfahr der Fledertiere fossil nicht bekannt ist, haben sich in der Fledermausforschung drei Hypothesen entwickelt: Die „Echoortung zuerst“, die „Flug zuerst“ und die „Tandem Hypothese“ (Flug und Echoortung entwickelten sich zusammen). Im Jahr 2020 ist noch eine vierte Hypothese hinzugekommen, die Hypothese der interdigitalen Flughäute (Flughäute zwischen den Vordergliedern).
Die „Echoortung zuerst“ Hypothese besagt, dass nachtaktive Fledermausvorfahren eine Greiftechnik zur Jagd von fliegenden Insekten verwendeten. Diese Technik beinhaltet das Ausstrecken der bereits verlängerten Vordergliedmaßen von einem Ast aus und ihr lag wahrscheinlich ein komplexes sensorisches System zugrunde zur Vorhersage von Beutebewegungen. Daraus entwickelte sich der Ultraschall, der erst zur Kommunikation untereinander verwendet wurde und sich dann in eine primitive Echoortung änderte, um so den Beutefang zu unterstützen.
Mit der Zeit entwickelte sich diese Echoortung immer mehr, die Vorderbeine verlängerten sich und eine Membran entstand zwischen den Gliedern, eine Art „Netz“ zum Beutefang. Später, durch Sprünge von Ast zu Ast, entwickelte sich erst der Gleitflug und dann der aktive Flug. Ein Punkt, der für die „Echoortung zuerst“ Hypothese spricht, ist die Tatsache, dass einige andere Säugetiere (vor allem die Spitzmäuse der Gattungen Sorex und Blarina) auch ohne Flugfähigkeit eine Echoortung entwickelt haben.
Die „Flug zuerst“ Hypothese besagt, dass die Vorfahren der Fledermäuse den Gleitflug entwickelt haben, während sie zwischen den Bäumen sprangen und dann der Gleitflug durch den aktiven Schlagflug ersetzt wurde, da dieser eine bessere Kontrolle und Manövrierfähigkeit ermöglichte. Die Echoortung war keine Voraussetzung für einen Flug und entwickelte sich erst mit dem Nachtleben der Fledermäuse. Da eine Echoortung zudem im Stand oder Laufen energetisch sehr aufwendig ist, aber im Flug weniger Energie hierfür verbraucht wird, muss die Flugfähigkeit vor der Evolution der Echoortung vorhanden gewesen sein. Der Fund der primitiven Fledermaus Onychonycteris, die schon aktiv fliegen konnte, aber noch keine Echoortung besaß, stützt die „Flug zuerst“ Hypothese.
Die Tandem Hypothese ist eine Konsequenz aus den ungeklärten Hypothesen „Echoortung zuerst“ und „Flug zuerst“. Diese Hypothese besagt, dass sich ein Echolokationssystem im Tandem mit dem Schlagflug entwickelt hat, so dass die Länge der Sprünge und damit der Beginn des ersten Gleitfluges zwischen den Ästen mit zunehmender Entwicklung der Echoortung zugenommen hat. Dies wird unterstützt durch die energetische Kopplung des aktiven Fluges an die Kehlkopfechoortung.
Die Hypothese der „interdigitalen Flughäute“ besagt, dass die Vorfahren der Fledermäuse
(Flug-)häute zwischen den Gliedmaßen vor der aktiven Flugfähigkeit entwickelten, dass sich der aktive Flug innerhalb der Fledertiere mehrfach unabhängig entwickelt haben könnte und dass die Ähnlichkeit der Flügelform in den verschiedenen Linien der Fledertiere durch Konvergenz (Ähnlichkeit, bedingt durch die gleiche Funktion) von einem gemeinsamen Vorfahren mit interdigitalen Flughäuten entstanden ist. Diese Hypothese ist eine Synthese aus dem aktuellen Verständnis des Fluges, der Evolution und der Phylogenie der Fledertiere.
Welche der Hypothesen letztendlich zutrifft oder ob alle Hypothesen falsch liegen, müssen zukünftige Fossilfunde zeigen.
Quellen
Anderson et al. 2020: The evolution of flight in bats: a novel hypothesis. Mammal Reviews 50: 426-439
Fenton et al. 1995: Signal strength, timing, and self-deafening: the evolution of echolocation in bats. Palaeobiology 21(2): 229-242
Gunnell et al. 2003: Oldest placental mammal from Sub-Saharan Africa: Eocene microbat from Tanzania – evidence for early evolution of sophisticated echolocation. Palaeontological Electronica 5(3)
Gunell und Simmons 2005: Fossil evidence and the origin of bats. Journal of Mammalian Evolution 12: 209-246
Habersetzer et al. 2008: Die Evolution des Fluges und der Echoortung. Biologie Unserer Zeit 4: 246-254
Simmons et al. 2008: Primitive early Eocene bat from Wyoming and the evolution of flight and echolocation. Nature 451: 818-822
Speakman 2001: The evolution of flight and echolocation in bats: another leap in the dark. Mammal Reviews 31: 111-130
Storch et al. 2002: Tachypteron franzeni n. gen., n. sp., earliest emballonurid bat from the middle Eocene of Messel (Mammalia, Chiroptera) Paläontologische Zeitschrift 76: 189-199
Teeling et al. 2005: A molecular phylogeny for bats illuminates biogeography and the fossil record. Science 307: 580-584
© Text, Foto und Zeichnungen Dr. Andreas Müller, Düsseldorf
Ein Gedanke zu „Millionen Jahre älter als der Mensch: Evolution der Fledertiere“
Kommentare sind geschlossen.